
Significance
RNA stem-loop folding is the process by which a single RNA strand bends back and pairs with itself to produce a short double-helical stem capped by an unpaired loop. It is one of the basic structural events that gives RNA its shape and much of its functional behavior. Complementary bases that are separated along the sequence have to find one another, pair in the correct register, and do so without forcing the intervening nucleotides into an unfavorable geometry. The unpaired segment connects the two sides of the stem and also affects how easily the structure forms, how stable it remains, and how readily it can reorganize. RNA function depends on its sequence, and also on what parts of the chain become paired, which bases stay exposed, how long a given structure survives, and how flexibly it can shift in response to proteins, ligands, ions, or changes in the cellular environment. Stem-loops appear throughout RNA biology. They are found in messenger RNAs, viral RNAs, ribozymes, riboswitches, and many regulatory noncoding RNAs. Sometimes they help protect a region from degradation. Sometimes they control translation or create a recognition surface for a protein. In larger RNAs, they often act as local organizing elements from which more complex structures can grow.
For that reason, stem-loop folding has drawn so much attention in simulation studies. These motifs are small enough to examine in atomic detail, but they are not simple in a physical sense. A model has to reproduce the balance among base pairing, stacking, backbone geometry, and electrostatic interactions well enough for the structure to emerge for the right reasons. That is what makes RNA stem-loop folding scientifically useful: it is both a real biological process and a demanding test of whether current molecular models are actually capturing how RNA structures form. For molecular dynamics, this makes stem-loops useful but rather unforgiving test cases. The stem mainly reports on the balance between base pairing and stacking, whereas the loop tests backbone geometry, noncanonical contacts, and local solvation effects. If stacking is too strong, misfolded compact structures may look artificially stable. If electrostatics or solvent response are treated too coarsely, loop and bulge geometries can move away from the experimental ensemble. Starting from extended chains therefore asks more than whether a model can preserve a known structure. It asks whether the simulation can recover, at least in part, the physical path by which an RNA fold comes into being.
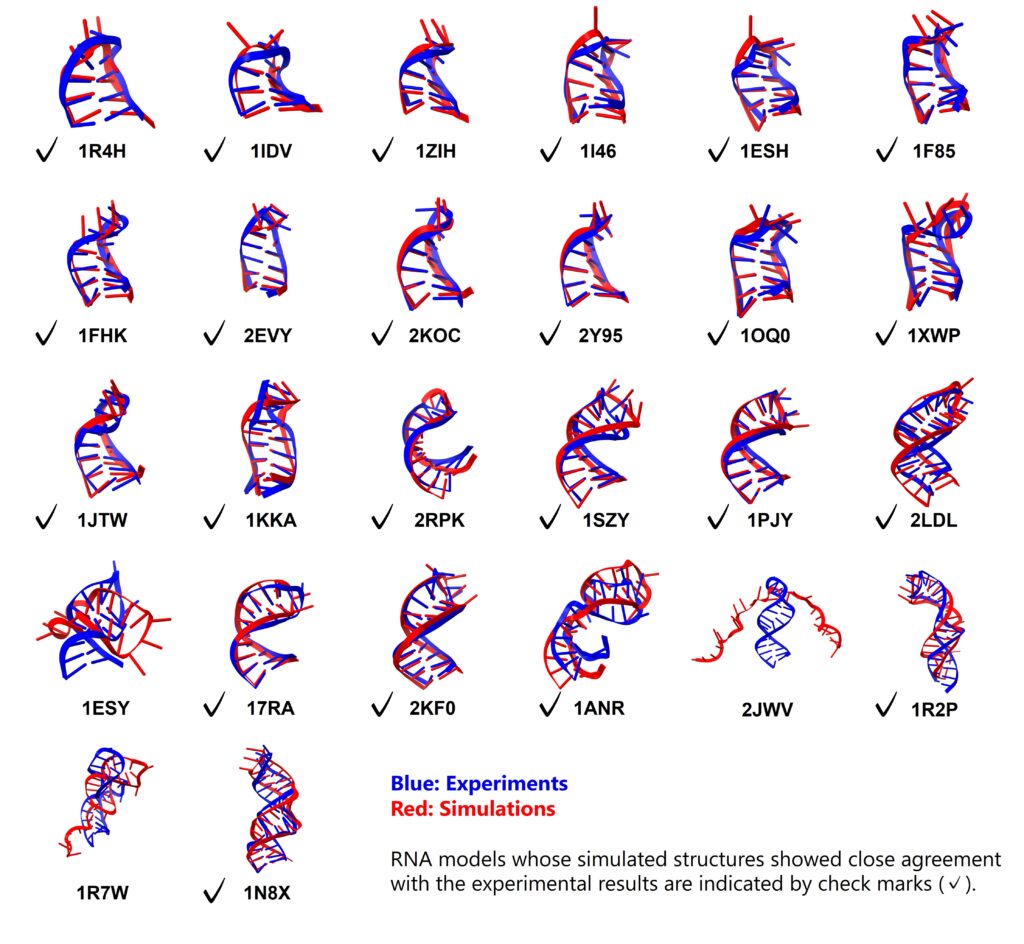
In a recent research paper published in ACS Omega, Professor Tadashi Ando from the Tokyo University of Science examined whether conventional molecular dynamics could fold RNA stem-loops from extended conformations using the DESRES-RNA force field and GB-neck2 implicit solvent. The simulations recovered native stem pairing in all simple stem-loops and in five of eight more complex models. The main technical advance is a benchmark that separates reliable stem-folding behavior from the remaining difficulty of loop and bulge modeling. Briefly, Professor Tadashi Ando performed de novo folding simulations on 26 RNA stem-loops, ranging from 10 to 36 nucleotides, including 18 simple stem-loops and eight structures containing bulges or internal loops. The investigators used conventional molecular dynamics at 298 K, with three independent trajectories for each model, and assessed folding through native base-pair recovery, RMSD values, clustering behavior, and comparison with experimentally determined NMR structures.
For the 18 stem-loops without bulges or internal loops, the authors observed folding into structures retaining all native stem base pairs. Most models reached stem-region RMSD values below 2 Å and whole-molecule RMSD values below 5 Å. The stems behaved as the most reliable structural element: once the correct base-pairing pattern formed, many trajectories maintained the folded state. The loop regions, however, remained less accurately described, with loop RMSD values near 4 Å in many cases. That separation between stem accuracy and loop imperfection is scientifically useful, because it identifies where the force-field and solvent approximation are working well and where local RNA chemistry remains more difficult to reproduce. For the eight stem-loops with bulges or internal loops, the researcher obtained complete native stem pairing in five cases. These more complex molecules often folded through a local route in which the stem adjoining the hairpin loop formed before the more terminal duplex region. The study also showed that several more difficult models sampled misaligned base-pairing arrangements. That behavior is scientifically informative because it reflects the kinetic cost of allowing nonnative contacts to become too stable in a reduced-solvent model.
Professor Tadashi Ando’s work demonstrated that stem formation, at least for many small RNA motifs, can now be recovered from extended conformations with impressive structural fidelity under a computationally efficient implicit-solvent protocol. Loop and bulge modeling still requires caution, especially when noncanonical hydrogen bonds, local base orientation, and solvent-specific contacts determine the experimental structure. These findings are important in RNA biology because many functional RNA interactions depend on single-stranded or partially paired regions, not only on ideal duplex stems. Riboswitches, RNA-protein interfaces, kissing-loop contacts, and ligand-binding pockets all place heavy demands on loop and bulge accuracy. Ando’s study therefore provides a practical benchmark for future simulations: success should be evaluated by whether the model can preserve the local chemistry that makes an RNA motif biologically recognizable. Another implication of the research work is that the DESRES-RNA and GB-neck2 combination which may be useful for exploring broad conformational searches, early folding events, and stem organization in RNA systems where explicit solvent sampling remains expensive. For detailed loop chemistry, explicit solvent treatment or further parameter refinement may still be needed.

Reference
Ando T. Molecular Dynamics Simulations of RNA Stem-Loop Folding Using an Atomistic Force Field and a Generalized Born Implicit Solvent. ACS Omega. 2025;10(43):51011-51027. doi: 10.1021/acsomega.5c05377.
Go to ACS Omega